Что такое днк и рнк? структура днк. функции днк
Содержание:
Транспортная рибонуклеиновая кислота
т-РНК наиболее изучены. Они составляют десять процентов клеточной рибонуклеиновой кислоты. Эти виды РНК связываются с аминокислотами благодаря специальному ферменту и доставляются на рибосомы. При этом аминокислоты переносятся транспортными молекулами. Однако бывает, что аминокислоту кодируют разные кодоны. Тогда переносить их будут несколько транспортных РНК.
Она сворачивается в клубочек, когда неактивна, а функционируя, имеет вид клеверного листа.
В ней различаются следующие участки:
- акцепторный стебель, имеющий последовательность нуклеотидов АЦЦ;
- участок, служащий для присоединения к рибосоме;
- антикодон, кодирующий аминокислоту, которая присоединена к этой т-РНК.
Строение и функции РНК
РНК — полимер, мономерами которой являются рибонуклеотиды. В отличие от ДНК, РНК образована не двумя, а одной полинуклеотидной цепочкой (исключение — некоторые РНК-содержащие вирусы имеют двухцепочечную РНК). Нуклеотиды РНК способны образовывать водородные связи между собой. Цепи РНК значительно короче цепей ДНК.
Мономер РНК — нуклеотид (рибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания РНК также относятся к классам пиримидинов и пуринов.
Пиримидиновые основания РНК — урацил, цитозин, пуриновые основания — аденин и гуанин. Моносахарид нуклеотида РНК представлен рибозой.
Выделяют три вида РНК: 1) информационная (матричная) РНК — иРНК (мРНК), 2) транспортная РНК — тРНК, 3) рибосомная РНК — рРНК.
Все виды РНК представляют собой неразветвленные полинуклеотиды, имеют специфическую пространственную конформацию и принимают участие в процессах синтеза белка. Информация о строении всех видов РНК хранится в ДНК. Процесс синтеза РНК на матрице ДНК называется транскрипцией.
Транспортные РНК содержат обычно 76 (от 75 до 95) нуклеотидов; молекулярная масса — 25 000–30 000. На долю тРНК приходится около 10% от общего содержания РНК в клетке. Функции тРНК: 1) транспорт аминокислот к месту синтеза белка, к рибосомам, 2) трансляционный посредник. В клетке встречается около 40 видов тРНК, каждый из них имеет характерную только для него последовательность нуклеотидов. Однако у всех тРНК имеется несколько внутримолекулярных комплементарных участков, из-за которых тРНК приобретают конформацию, напоминающую по форме лист клевера. У любой тРНК есть петля для контакта с рибосомой (1), антикодоновая петля (2), петля для контакта с ферментом (3), акцепторный стебель (4), антикодон (5). Аминокислота присоединяется к 3′-концу акцепторного стебля. Антикодон — три нуклеотида, «опознающие» кодон иРНК. Следует подчеркнуть, что конкретная тРНК может транспортировать строго определенную аминокислоту, соответствующую ее антикодону. Специфичность соединения аминокислоты и тРНК достигается благодаря свойствам фермента аминоацил-тРНК-синтетаза.
Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000. На долю рРНК приходится 80–85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы — органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках. Функции рРНК: 1) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом; 2) обеспечение взаимодействия рибосомы и тРНК; 3) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания, 4) формирование активного центра рибосомы.
Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от 50 000 до 4 000 000). На долю иРНК приходится до 5% от общего содержания РНК в клетке. Функции иРНК: 1) перенос генетической информации от ДНК к рибосомам, 2) матрица для синтеза молекулы белка, 3) определение аминокислотной последовательности первичной структуры белковой молекулы.
Синтез
Транскрипция РНК из ДНК с участием фермента РНК-полимеразы II
Основные статьи: Транскрипция (биология), Редактирование РНК, Сплайсинг
Синтез РНК в живой клетке проводится ферментом — РНК-полимеразой. У эукариот разные типы РНК синтезируются разными, специализированными РНК-полимеразами. В целом матрицей синтеза РНК может выступать как ДНК, так и другая молекула РНК. Например, полиовирусы используют РНК-зависимую РНК-полимеразу для репликации своего генетического материала, состоящего из РНК. Но РНК-зависимый синтез РНК, который раньше считался характерным только для вирусов, происходит и в клеточных организмах, в процессе так называемой РНК-интерференции.
Как в случае ДНК-зависимой РНК-полимеразы, так и в случае РНК-зависимой РНК-полимеразы фермент присоединяется к промоторной последовательности. Вторичная структура молекулы матрицы расплетается с помощью хеликазной активности полимеразы, которая при движении субстрата в направлении от 3′ к 5′ концу молекулы синтезирует РНК в направлении 5′ → 3′. Терминатор транскрипции в исходной молекуле определяет окончание синтеза. Многие молекулы РНК синтезируются в качестве молекул-предшественников, которые подвергаются «редактированию» — удалению ненужных частей с помощью РНК-белковых комплексов.
Например, у кишечной палочки гены рРНК расположены в составе одного оперона (в rrnB порядок расположения такой: 16S — tRNAGlu2 — 23S —5S) считываются в виде одной длинной молекулы, которая затем подвергается расщеплению в нескольких участках с образованием сначала пре-рРНК, а затем зрелых молекул рРНК. Процесс изменения нуклеотидной последовательности РНК после синтеза носит название процессинга или редактирования РНК.
После завершения транскрипции РНК часто подвергается модификациям (см. выше), которые зависят от функции, выполняемой данной молекулой. У эукариот процесс «созревания» РНК, то есть её подготовки к синтезу белка, часто включает сплайсинг: удаление некодирующих белок последовательностей (интронов) с помощью рибонуклеопротеида сплайсосомы. Затем к 5′ концу молекулы пре-мРНК эукариот добавляется особый модифицированный нуклеотид (кэп), а к 3′ концу несколько аденинов, так называемый «полиА-хвост».
Игрок № 1 — короткие интерферирующие РНК
Функции киРНК в клетках беспозвоночных достаточно разнообразны. Первая и основная — это иммунная защита. «Традиционная» иммунная система (лимфоциты + лейкоциты + макрофаги) присутствует лишь у сложных многоклеточных организмов. У одноклеточных же, беспозвоночных и растений (у которых такой системы либо нет, либо она находится в зачаточном состоянии) иммунная защита строится на основе РНК-интерференции. Иммунитет, основанный на РНК-интерференции, не нуждается в сложных органах «тренировки» предшественников иммунных клеток (селезенка, тимус); в то же время, многообразие теоретически возможных последовательностей коротких РНК (421 вариантов) соотносимо с числом возможных белковых антител высших животных. Кроме того, киРНК синтезируются на основе инфицировавшей клетку «враждебной» РНК, а значит, в отличие от антител, они сразу «затачиваются» под конкретный тип инфекции. И хотя вне клетки защита на основе РНК-интерференции не работает (по крайней мере, таких данных пока нет), внутриклеточный иммунитет она обеспечивает более чем удовлетворительно.
Прежде всего, киРНК создаёт антивирусный иммунитет, уничтожая мРНК или геномную РНК инфекционных организмов (например, так киРНК и были открыты у растений ). Введение вирусной РНК вызывает мощную амплификацию специфических киРНК на основе молекулы-затравки — самой вирусной РНК. Кроме того, киРНК подавляют экспрессию различных мобильных генетических элементов (МГЭ), а значит, обеспечивает защиту и от эндогенных «инфекций». Мутации в генах RISC-комплекса часто ведут к повышению нестабильности генома из-за высокой активности МГЭ; киРНК может быть ограничителем экспрессии собственных генов, срабатывая в ответ на их гиперэкспрессию. Регуляция работы генов может происходить не только на уровне трансляции, но и во время транскрипции — через метилирование генов по гистону Н3.
В современной экспериментальной биологии значение РНК-интерференции и коротких РНК трудно переоценить. Разработана технология «выключения» (или нокдауна) отдельных генов in vitro (на культурах клеток) и in vivo (на эмбрионах), что уже стало стандартом de facto при изучении любого гена. Иногда даже, чтобы установить роль отдельных генов в каком-нибудь процессе, проводят систематическое «выключение» всех генов по очереди .
Возможностью применения киРНК заинтересовались и фармацевты, поскольку способность направленной регуляции работы отдельных генов сулит неслыханные перспективы в лечении массы заболеваний. Небольшой размер и высокая специфичность действия обещают высокую эффективность и низкую токсичность лекарств на основе киРНК; однако решить проблему доставки киРНК к больным клеткам в организме пока не удалось — виной тому хрупкость и недолговечность этих молекул. И хотя сейчас десятки коллективов пытаются найти способ направлять эти «волшебные пули» точно в цель (внутрь больных органов), видимых успехов они пока не достигли. Кроме этого, есть и другие сложности. Например, в случае антивирусной терапии высокая избирательность действия киРНК может оказать «медвежью услугу» — поскольку вирусы быстро мутируют, изменённый штамм очень быстро потеряет чувствительность к киРНК, подобранной в начале терапии: известно, что замена всего лишь одного нуклеотида в киРНК приводит к существенному снижению эффекта интерференции.
В этом месте стоит напомнить ещё раз — киРНК были обнаружены только у растений, беспозвоночных и одноклеточных; хотя гомологи белков для РНК-интерференции (Dicer, RISC-комплекс) присутствуют и у высших животных, киРНК привычными методами не обнаруживались. Каково же было удивление, когда искусственно введённые синтетические аналоги киРНК вызывали сильный специфический дозозависимый эффект в культурах клеток млекопитающих! Это означало, что в клетках позвоночных РНК-интерференция не заместилась более сложными системами иммунитета, а эволюционировала вместе с организмами, превратившись во что-то более «продвинутое». Следовательно, у млекопитающих надо было искать не точные аналоги киРНК, а их эволюционных преемников.
Генотипирование вируса гепатита С
Гепатит С — упрощенное название целого спектра вирусов, которые сгруппированы по генотипам и субтипам по различиям в структуре РНК. Соответственно, реакции на воздействие препаратов будут индивидуальными.
Из 11 известных генотипов наибольшее распространение в мире получили 6. Субтипов насчитывается около 500, и они отличаются своеобразной чувствительностью к препаратам.
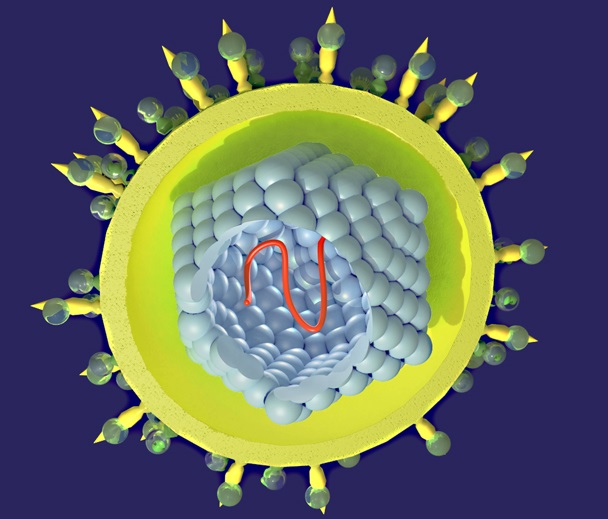
Анализ на генотип вируса С нужен, чтобы подобрать адекватное лечение для каждого конкретного случая заболевания. От схемы терапии зависит ее продолжительность и результативность.
Итоги исследования позволяют спрогнозировать развитие заболевания, подобрать приемлемые терапевтические мероприятия, дозировки препаратов. В некоторых случаях и биопсия печени проводится только после выполнения генотипирования.
Генотипы 1 – 6
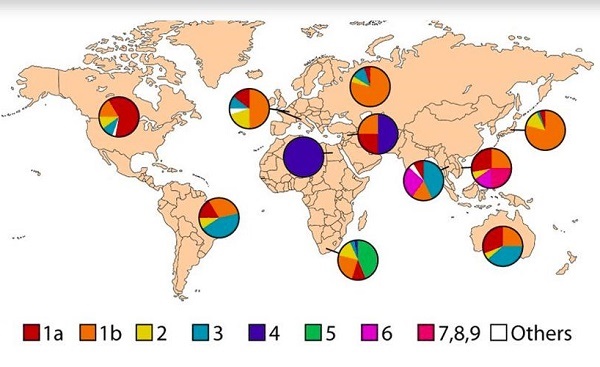
Генотипы возбудителя распределены по миру не одинаково:
- генотипы 1, 2, 3 типы наиболее распространены во всем мире;
- 1 и 2 имеют перевес в Западной Европе и Дальнем Востоке;
- 1 генотип – США;
- 4й –Северная Африка;
- 6й – Южная Африка;
- 1В тип – Япония.
Для постсоветского пространства характерными являются генотипы 1, 2 и 3 вируса гепатита С.
Генотипы 1b
Из субтипов на территории Центральной и Восточной Европы, а также в Азии наиболее часто встречается вирус гепатита С 1b. Его специфика:
- Форма недуга в основном хроническая.
- Бессимптомное протекание болезни (пациент может узнать о своей проблеме через десятилетия после заражения).
Вирус с высокой долей вероятности спровоцирует цирроз печени, гепатоцеллюлярную карциному, внепеченочные осложнения ( криоглобулинемический васкулит , злокачественные опухоли лимфатической системы), которые могут привести к летальному исходу.
Интерфероновые схемы лечения практически не дают никакой реакции. Терапия разновидности Даклатасвир + Асунапревир/Софосбувир позволяет достичь стойкого вирусологического ответа.
Генотипы 3а и 3в
Следующими по распространенности на территории Украины, Белоруссии и России являются РНК вируса гепатита С 3а и 3в. Он:
- значительно реже протекает в хронической форме;
- характеризуется поражением желчевыводящих путей и стеатозом (накоплением жира в печеночных клетках);
- реже приводит к циррозу;
- при подборе дозировки Рибавирина следует отталкиваться от веса пациента, а для недуга генотипа 3a количество препарата назначается врачом.
История изучения
Нуклеиновые кислоты были открыты в 1868 году швейцарским учёным Иоганном Фридрихом Мишером, который назвал эти вещества «нуклеин», поскольку они были обнаружены в ядре (лат. nucleus). Позже было обнаружено, что бактериальные клетки, в которых нет ядра, тоже содержат нуклеиновые кислоты. Значение РНК в синтезе белков было предположено в 1939 году в работе Торбьёрна Оскара Касперссона, Жана Брачета и Джека Шульца. Джерард Маирбакс выделил первую матричную РНК, кодирующую гемоглобин кролика и показал, что при её введении в ооциты образуется тот же самый белок. В 1956—1957 годах А. Белозёрским, А. Спириным, Э. Волкиным, Л. Астраханом проводились работы по определению состава РНК клеток, которые привели к выводу, что основную массу РНК в клетке составляет рибосомальная РНК. Северо Очоа получил Нобелевскую премию по медицине в 1959 году за открытие механизма синтеза РНК. Последовательность 77 нуклеотидов одной из тРНК дрожжей S. cerevisiae была определена в 1965 году в лаборатории Роберта Холея, за что в 1968 году он получил Нобелевскую премию по медицине. В 1967 Карл Вёзе предположил, что РНК обладают каталитическими свойствами. Он выдвинул так называемую гипотезу мира РНК, в котором РНК прото-организмов служила и в качестве молекулы хранения информации (сейчас эта роль выполняется в основном ДНК) и молекулы, которая катализировала метаболические реакции (сейчас это делают в основном ферменты). В 1976 Уолтер Фаэрс и его группа в Гентском Университете в Бельгии определили первую последовательность генома РНК-содержащего вируса, бактериофага MS2. В начале 1990-х было обнаружено, что введение чужеродных генов в геном растений приводит к подавлению выражения аналогичных генов растения. Приблизительно в это же время было показано, что РНК длиной около 22 оснований, которые сейчас называются микроРНК, играют регуляторную роль в онтогенезе нематод C. elegans.
Функции
Первоначально Y-РНК были описаны как некодирующие РНК, связывающиеся с белком Ro60, который является антигеном, распознаваемым антителами из крови пациентов с системной красной волчанкой и синдромом Шегрена. Ro60 у позвоночных консервативен, его гомологи были выявлены у большинства Metazoa, а также в 5 % бактериальных геномов (среди которых геномы D. radiodurans и Salmonella). У всех этих организмов гомологи Ro60 связываются с Y-РНК или подобными им РНК с образованием рибонуклеопротеинов. У позвоночных Ro60 взаимодействует с нижним стеблевым доменом Y-РНК. С частью рибонуклеопротеинов, состоящих из Ro60 и Y-РНК (RoRNP), связывается белок La. Он необходим для правильной терминации транскрипции, опосредованной РНК-полимеразой III, и связывается с 3′-концевым хвостом новосинтезированных РНК в ядре. Большинство РНК впоследствии утрачивают полиуридиновый хвост, но Y-РНК его сохраняют и продолжают взаимодействовать с La. Он участвует в накоплении Y-РНК в ядре и защищает их от разрушения экзонуклеазами.
RoRNP участвуют в контроле качества некодирующих РНК, регулируют стабильность РНК и у некоторых видов участвуют в клеточном ответе на стресс. У многих видов, в числе которых лягушка Xenopus laevis, нематода C. elegans и домовая мышь, Ro60 связывается с такими дефектными некодирующими РНК, как неправильно уложенные 5S рРНК и . Нематоды, лишённые гомолога Ro60, жизнеспособны, однако у них нарушается процесс формирования особой личинки, предназначенной для перенесения неблагоприятных условий. Под действием УФ-излучения в клетках мыши и D. radiodurans происходит накопление RoRNP, и в отсутствие Ro60 их устойчивость к УФ-излучению понижается. У D. radiodurans ортолог Ro60 участвует в созревании рРНК, экспрессия которых была индуцирована тепловым стрессом, и разрушении рРНК, вызванном голоданием. Связывание Ro60 с неправильно уложенными некодирующими РНК неспецифично в отношении последовательности, и, по-видимому, этот белок может связываться с самыми разными РНК. По этой причине RoRNP можно рассматривать как внутриклеточные сенсоры стресса. В отсутствие Ro60 уровень Y-РНК в клетках как эукариот, так и прокариот снижался, из чего, вероятно, следует, что Ro60 стабилизирует Y-РНК, с которыми он взаимодействует.
Y-РНК участвует в инициации репликации хромосомной ДНК, а именно в формировании новых репликативных вилок. Однако они не нужны для элонгации и репликации и самого процесса удвоения ДНК как такового. Y-РНК вовлечены в инициацию репликации ДНК не сами по себе, а посредством белков. Для участия в инициации репликации ДНК необходим верхний стебель Y-РНК, но не участки связывания с белками Ro60 и La и не сами эти белки. Фактически около половины Y-РНК в человеческих клетках существует вне комплексов с белками Ro60 и La. Нарушение работы Y-РНК с помощью РНК-интерференции блокирует репликацию ДНК и пролиферацию клеток млекопитающих. При этом нормальный фенотип возвращается при введении в такие клетки короткой двуцепочечной РНК, соответствующей верхнему стеблю РНК hY1. Инактивация Y-РНК с помощью морфолиновых олигонуклеотидов в эмбрионах X. laevis и рыбы Danio rerio приводит к остановке развития и гибели на ранних этапах эмбриогенеза. В человеческих опухолях, напротив, наблюдается сверхэкспрессия Y-РНК по сравнению с нормальными тканями. Последовательности верхних стеблей Y-РНК таких организмов, как C. elegans, B. floridae и D. radiodurans, сильно отличаются от последовательностей позвоночных, и эти Y-РНК не могут функционально заменить Y-РНК в клетках человека.
Конкретный механизм участия Y-РНК в инициации репликации ДНК неизвестен. Известно, однако, что Y-РНК взаимодействует с несколькими белками, вовлечёнными в инициацию репликации ДНК, в числе которых репликации ORC, а также , и DUE-B, но не взаимодействует с белками репликативной вилки. С помощью Y-РНК удалось показать, что у человека Y-РНК динамически взаимодействуют с нереплицированным хроматином в G1-фазе клеточного цикла, колокализуясь с несколькими белками инициации репликации ДНК. После того как репликация началась, Y-РНК вытесняются из места начала репликации и не выявляются в местах активного синтеза ДНК.
Информационная РНК (иРНК)
Почти все РНК синтезируются на ДНК в процессе транскрипции. Однако часто транскрипция упоминается как синтез именно информационной РНК (иРНК). Связано это с тем, что последовательность нуклеотидов иРНК в последствии определит последовательность аминокислот синтезируемого в процессе трансляции белка.
Перед транскрипцией нити ДНК расплетаются, и на одной из них с помощью комплекса белков-ферментов синтезируется РНК по принципу комплементарности, так же как это происходит при репликации ДНК. Только напротив аденина ДНК к молекуле РНК присоединяется нуклеотид, содержащий урацил, а не тимин.
На самом деле на ДНК синтезируется не готовая информационная РНК, а ее предшественник — пре-иРНК. Предшественник содержит участки последовательности нуклеотидов, которые не кодируют белок и которые после синтеза пре-иРНК вырезаются при участии малых ядерных и ядрышковых РНК («дополнительные» типы РНК). Эти удаляющиеся участки называются интронами. Остающиеся части иРНК называются экзонами. После удаления интронов экзоны сшиваются между собой. Процесс удаления интронов и сшивания экзонов называется сплайсингом. Усложняющей жизнь особенностью является то, что можно вырезать интроны по-разному, в результате получатся разные готовые иРНК, которые будут служить матрицами для разных белков. Таким образом, вроде бы один ген ДНК может играть роль нескольких генов.
Следует отметить, что у прокариотических организмов сплайсинга не происходит. Обычно их иРНК сразу после синтеза на ДНК готова к трансляции. Бывает, что пока конец молекулы иРНК еще транскрибируется, на ее начале уже сидят рибосомы, синтезирующие белок.
После того как пре-иРНК созревает в информационную РНК и оказывается вне ядра, она становится матрицей для синтеза полипептида. При этом на нее «насаживаются» рибосомы (не сразу, какая-то оказывается первой, другая — второй и т. д.). Каждая синтезирует свою копию белка, т. е. на одной молекуле РНК могут синтезироваться сразу несколько одинаковых белковых молекул (понятно, что каждая будет находиться на своей стадии синтеза).
Рибосома, передвигаясь от начала иРНК к ее концу, считывает по три нуклеотида (хотя вмещает шесть, т. е. два кодона) и присоединяет соответствующую транспортную РНК (имеющую соответствующий кодону антикодон), к которой присоединена соответствующая аминокислота. После этого с помощью активного центра рибосомы ранее синтезированная часть полипептида, соединенная с предшествующей тРНК, как-бы «пересаживается» (образуется пептидная связь) на аминокислоту, прикрепленную к только что пришедшей тРНК. Таким образом, молекула белка постепенно увеличивается.
Когда молекула информационной РНК становится не нужна, клетка ее разрушает.
Действие РНК-полимеразы
Связывание и инициирование транскрипции
Схема инициализации транскрипции
В связывании РНК-полимеразы участвует α-субъединица, распознающая элемент ДНК, предшествующий гену (-40…-70 шагов), и σ-фактор, распознающий участок −10…-35. Существует большое количество σ-факторов, контролирующих экспрессию генов. Например: σ70, который синтезируется в нормальных условиях и позволяет РНК-полимеразе связываться с генами, отвечающими за метаболические процессы клетки; или σ32, блокирующий связывание РНК-полимеразы с генами белков теплового шока.
После связывания с ДНК структура РНК-полимеразы превращается из закрытой в открытую. Это превращение включает в себя разделение моноспиралей ДНК с образованием раскрученного участка длиной около 13 шагов. Рибонуклеотиды затем собираются в цепочку в соответствии с базовой нитью ДНК, используемой в качестве шаблона. Суперскрученность молекул ДНК играет существенную роль в деятельности РНК-полимеразы: поскольку участок ДНК перед РНК-полимеразой раскручен, в нем существуют положительные компенсационные супервитки. Участки ДНК позади РНК-полимеразы снова закручиваются и в них присутствуют отрицательные супервитки.
Элонгация
Во время элонгационной фазы транскрипции происходит добавление рибонуклеотидов к цепи и переход от структуры РНК-полимеразного комплекса от открытой к транскрипционной. По мере сборки молекулы РНК участок ДНК перед РНК-полимеразой раскручивается далее, и 13-парный открытый комплекс превращается в 17-парный транскрипционный комплекс. В этот момент промотор (участок ДНК −10…-35 шагов) завершается, и σ-фактор отделяется от РНК-полимеразы. Это позволяет остальному РНК-полимеразному комплексу начать движение вперед, так как σ-фактор удерживал его на месте.
17-парный транскрипционный комплекс содержит гибрид ДНК и РНК, содержащий 8 пар оснований — 8-шаговый участок РНК, соединенный с шаблонной цепью ДНК. По мере выполнения транскрипции рибонуклеотиды добавляются к 3′-концу собираемой РНК, и РНК-полимеразный комплекс движется по цепи ДНК. Хотя в РНК-полимеразе не обнаружено свойств, характерных для 3′-экзонуклеазы, аналогичных проверочной деятельности ДНК-полимеразы, есть свидетельства того, что РНК-полимераза останавливается и корректирует ошибки в случаях ошибочного формирования пар оснований ДНК-РНК.
Добавление рибонуклеотидов к РНК обладает механизмом, очень близким к полимеризации ДНК. Считается, что ДНК- и РНК-полимеразы могут быть эволюционно связаны. Аспарагиновые остатки в РНК-полимеразе связываются с ионами Mg2+, которые, в свою очередь, осуществляют выравнивание фосфатных групп рибонуклеотидов: первый Mg2+ удерживает α-фосфат нуклеотидтрифосфата, подлежащего добавлению в цепочку. Это позволяет осуществить связывание нуклеотида с 3′ OH-группой конца собираемой цепочки и таким образом добавить НТФ в цепочку. Второй Mg2+ удерживает пирофосфат НТФ. Общее уравнение реакции таким образом имеет вид:
(НМФ)n + НТФ —> (НМФ)n+1 + ПФi
Терминация
Терминация транскрипции РНК может быть ρ-независимой либо ρ-зависимой.
ρ-независимая терминация осуществляется без помощи ρ-фактора. Транскрипция палиндромного участка ДНК приводит к формированию шпильки из РНК, зацикленной и связанной с самой собой. Эта шпилька богата гуанином и цитозином, что делает её более стабильной, нежели гибрид ДНК-РНК. В результате 8-парный гибрид ДНК-РНК в транскрипционном комплексе сокращается до 4-парного. В случае если эти 4 последние пары оснований составлены слабыми аденином и уридином, молекула РНК отделяется.
Характеристики
Одноцепочечные РНК-вирусы и смысловая РНК
РНК-вирусы могут быть дополнительно классифицированы в соответствии с полярностью их РНК на и , или РНК . Вирусы, содержащие одноцепочечную (+)РНК похожую на мРНК и таким образом может непосредственно транслирована зараженной клеткой. Поэтому (+)РНК также называется смысловой. Антисмысловая (-)РНК вируса комплиментарна мРНК и таким образом может быть превращена в (+)РНК действием РНК-зависимой РНК-полимеразой перед трансляцией. Очищенная РНК (+)РНК-содержащих вирусов сама по себе может напрямую вызывать инфекцию, хотя ее инфекционность ниже, чем у полноценных вирусных частиц. Очищенная РНК (-)РНК-содержащих вирусов неинфекционна сама по себе, так как она должна транскрибироваться в (+)РНК; каждый вирион после транскрипции может быть источником нескольких смысловых цепей (+)РНК. Двухцепочечные(±)РНК вирусы напоминают (-)РНК-содержащие вирусы тем, что они также транслируют гены с отрицательной (некодирующей) цепи РНК.
Двуцепочечные РНК вирусы
Структура вириона реовируса
Двуцепочечные РНК вирусы представляет разнообразную группу вирусов отличающихся по заражаемому кругу хозяев (людей, животных, растений, грибов, и бактерий), числу фрагментов генома (от 1 до 12 сегментов), и организацией вириона (формой капсида, количеством слоев капсида, выросты, шипы, и т.п.). Члены этой группы включают ротавирусы, всемирно известные как наиболее распространенная причина детских гастроэнтеритов, и пикорнавирусы, известный во всем мире как наиболее часто встречающийся вирус в фекалиях людей и животных с признаками диареи или без них. является экономически важным возбудителем болезни коз и овец. В прошлые годы, достигнут большой прогресс в определении атомной и субнанометровой структуры ряда ключевых вирусных белков и капсидов вирионов нескольких дцРНК-вирусов, подчеркивая существенные параллели в структуре и механизмах процессов репликации многих из этих вирусов.
Частота мутаций
РНК вирусы имеют очень высокую частоту мутаций по-сравнению с ДНК-вирусами, потому что вирусная РНК-зависимая РНК-полимераза не имеет способности (3′-5′-экзонуклеазной гидролитической активности) к коррекции ошибок, как ДНК-полимеразы. Это одна из причин того, почему так трудно сделать эффективную вакцину для предотвращения болезней, вызываемых РНК-вирусами.
Ретровирусы также имеют высокую частоту мутаций, даже несмотря на то, что их промежуточная форма генома в виде ДНК интегрируется в геном хозяина (и, следовательно, после интеграции в геном, во время репликации, подвергается корректирующей проверке ДНК-полимеразой хозяина). Высокая частота мутаций обусловлена высокой частотой ошибок во время обратной транскрипции, до стадии интеграции в геном хозяина. Ошибки при этом встраиваются в обе цепи ДНК.
Некоторые гены РНК-вирусов важны для их репликации и мутации в них не допустимы. Например, участок генома вируса гепатита С кодирующий коровый белок, высоко консервативен, потому что он содержит структурную РНК, вовлеченную в образование участка внутренней посадки рибосомы.
Группа IV — оц(+)РНК-вирусы
Основная статья: Одноцепочечные РНК-вирусы с позитивной цепью
В этой группе есть три порядка и 34 семейства. Кроме того, существует ряд неклассифицированных и родов.
- Порядок Nidovirales
- Семейство Arteriviridae
- Семейство Coronaviridae — включает Coronavirus, SARS, MERS
- Семейство Mesoniviridae
- Семейство Roniviridae
- Порядок Picornavirales
- Семейство Dicistroviridae
- Семейство Iflaviridae
- Семейство Marnaviridae
- Семейство Picornaviridae — включает Poliovirus, Rhinovirus (общая простуда), вирус гепатита А
- Семейство Secoviridae включает подсемейство Comovirinae
- Род Bacillariornavirus
- Род Dicipivirus
- Род Labyrnavirus
- Род Sequiviridae
- Вид Kelp fly virus
- Порядок Tymovirales
- Семейство Alphaflexiviridae
- Семейство Betaflexiviridae
- Семейство Gammaflexiviridae
- Семейство Tymoviridae
- Неклассифицированные
- Семейство Alphatetraviridae
- Семейство Alvernaviridae
- Семейство Astroviridae
- Семейство Barnaviridae
- Семейство Benyviridae
- Семейство Bromoviridae
- Семейство Caliciviridae — включает Norovirus
- Семейство Carmotetraviridae
- Семейство Closteroviridae
- Семейство Flaviviridae — включает вирус жёлтой лихорадки, вирус лихорадки Западного Нила, вирус гепатита С, вирус лихорадки денге, вирус Зика
- Семейство Fusariviridae
- Семейство Hepeviridae
- Семейство Hypoviridae
- Семейство Leviviridae
- Семейство Luteoviridae — включает Barley yellow dwarf virus
- Семейство Polycipiviridae
- Семейство Narnaviridae
- Семейство Nodaviridae
- Семейство Permutotetraviridae
- Семейство Potyviridae
- Семейство Sarthroviridae
- Семейство Statovirus
- Семейство Togaviridae — включает вирус краснухи, Ross River virus, Sindbis virus, Вирус Чикунгунья
- Семейство Tombusviridae
- Семейство Virgaviridae
- Неклассифицированные роды
- Род Blunervirus
- Род Cilevirus
- Род Higrevirus
- Род Idaeovirus
- Род Negevirus
- Род Ourmiavirus
- Род Polemovirus
- Род Sinaivirus
- Род Sobemovirus
- Неклассифицированные виды
- Acyrthosiphon pisum virus
- Bastrovirus
- Blackford virus
- Blueberry necrotic ring blotch virus
- Cadicistrovirus
- Chara australis virus
- Extra small virus
- Goji berry chlorosis virus
- Hepelivirus
- Jingmen tick virus
- Le Blanc virus
- Nedicistrovirus
- Nesidiocoris tenuis virus 1
- Niflavirus
- Nylanderia fulva virus 1
- Orsay virus
- Osedax japonicus RNA virus 1
- Picalivirus
- Plasmopara halstedii virus
- Rosellinia necatrix fusarivirus 1
- Santeuil virus
- Secalivirus
- Solenopsis invicta virus 3
- Wuhan large pig roundworm virus
Вирусы-сателлиты
- Семейство Sarthroviridae
- Род Albetovirus
- Род Aumaivirus
- Род Papanivirus
- Род Virtovirus
- Chronic bee paralysis virus
Также был описан неклассифицированный астровирусный / гепевирусоподобный вирус.
Управление транскрипцией
Электронная микрофотография нитей ДНК, обвешанных сотнями молекул РНК-полимеразы, слишком маленьких для такого разрешения. Каждая РНК-полимераза транскрибирует нить РНК, которая видна на фотографии как ответвление от ДНК. Отметкой «Begin» указан 5′-конец ДНК, с которого РНК-полимераза начинает транскрипцию; «End» — 3′-конец, у которого транскрипция более длинных молекул РНК завершается.
Управление процессом транскрипции генов позволяет контролировать экспрессию генов и таким образом позволяет клетке адаптироваться к изменяющимся условиям внешней среды, поддерживать метаболические процессы на должном уровне, а также выполнять специфические функции, необходимые для существования организма. Неудивительно, что действие РНК-полимеразы очень сложно и зависит от множества факторов (так, у Escherichia coli идентифицировано более 100 факторов, тем или иным способом влияющих на РНК-полимеразу).
РНК-полимераза начинает транскрипцию с особых участков ДНК, называемых промоторами и производит цепочку РНК, комплементарную соответствующей части нити ДНК.
Процесс наращивания молекулы РНК нуклеотидами называется элонгацией. В эукариотических клетках РНК-полимераза может собирать цепочки из более 2,4 млн элементов (например, такую длину имеет полный ген белка дистрофина).
РНК-полимераза завершает формирование цепочки РНК, когда встречает в ДНК специфическую последовательность, называемую терминатором.
РНК-полимераза производит следующие разновидности РНК:
- Матричная РНК (мРНК) — шаблон для синтеза белков в рибосомах.
-
Некодирующая РНК или «РНК-ген» — большой класс генов, кодирующих РНК, на которых не может быть построено белка. Самые известные представители этого класса — транспортная РНК (тРНК) и рибосомная РНК (рРНК), сами участвующие в процессе синтеза белка. Однако начиная с поздних 90-х годов XX столетия было обнаружено много других РНК-генов. Это дало возможность предположить, что РНК-гены играют более значительную роль в клетке, чем было принято считать раньше.
- Транспортная РНК (тРНК), переносящая аминокислоты к растущей на рибосоме белковой цепочке во время процесса трансляции.
- Рибосомная РНК (рРНК), входящая в состав рибосомы;
- МикроРНК, регулирующая активность генов;
- Каталитическая РНК, обладающая свойствами ферментов.
РНК-полимераза осуществляет синтез с нуля. Это возможно вследствие того, что взаимодействие начального нуклеотида гена и РНК-полимеразы позволяет ей закрепиться на цепочке и обрабатывать следующие нуклеотиды. Это отчасти объясняет, почему РНК-полимераза обычно начинает транксрипцию с АТФ, за которым следует ГТФ, УТФ и затем ЦТФ.
В отличие от ДНК-полимеразы РНК-полимераза обладает также геликазным действием.
Что такое ДНК?
ДНК расшифровывается как дезоксирибонуклеиновая кислота. Она представляет собой одну из трех макромолекул клетки (две другие – белки и рибонуклеиновая кислота), которая обеспечивает сохранение и передачу генетического кода развития и деятельности организмов. Простыми словами, ДНК – носитель генетической информации. В ее составе содержится генотип индивида, который обладает способностью к самовоспроизводству и передает информацию по наследству.
Как химическое вещество кислота была выделена из клеток еще в 1860-х годах, однако вплоть до середины XX столетия никто и не предполагал, что она способна хранить и передавать информацию. Долгое время считалось, что эти функции выполняют белки, однако в 1953 году группа биологов сумела значительно расширить понимание сути молекулы и доказать первостепенную роль ДНК в сохранении и передаче генотипа. Находка стала открытием века, а ученые получили за свою работу Нобелевскую премию.
Долгое время считалось, что эти функции выполняют белки, однако в 1953 году группа биологов сумела значительно расширить понимание сути молекулы и доказать первостепенную роль ДНК в сохранении и передаче генотипа. Находка стала открытием века, а ученые получили за свою работу Нобелевскую премию.